您当前的位置:检测资讯 > 科研开发
嘉峪检测网 2024-08-30 20:48
摘 要 / Abstract
麦角硫因(EGT)是一种天然的小分子组氨酸硫脲衍生物,存在于微生物细胞和动植物体内。EGT可由真菌、蓝细菌、放线菌等微生物合成,人体自身不能合成EGT,需通过饮食摄取,食用菌是EGT的重要饮食来源之一。EGT具有抗氧化、抗炎、免疫调节等多种生物学功能,在食品、药品和化妆品等领域具有广泛的应用。本文综述了EGT的生物学功能、制备方法及其应用的研究进展。
Ergothioneine (EGT) is a natural small molecule histidine thiourea derivative which exists in microbial cells,animals and plants.EGT can be synthesized by microorganisms such as fungi,cyanobacteria,and actinomycetes.The human body cannot synthesize EGT itself,and must obtain it through dietary intake.Edible fungi are one of the important dietary sources of EGT.EGT exhibits various biological functions such as anti-oxidation,anti-inflammation and immune regulation,and is widely used in the fields of food,drugs and cosmetics.This paper reviews the biological functions,preparation methods and applications of EGT.
关 键 词 / Key words
麦角硫因;生物学功能;抗氧化;生物合成;应用
ergothioneine; biological function; anti-oxidation; biosynthesis; application
麦角硫因(ergothioneine,EGT)是一种天然的含硫氨基酸,最初于1909年由Charles Tanret在麦角菌(Claviceps purpurea)中分离得到,并以该真菌的名称命名[1]。EGT广泛存在于微生物细胞、动植物体内,在蘑菇、黑豆、红豆、燕麦等食物中含量较高[2]。EGT也存在于人体各种细胞和组织中,其中红细胞、骨髓、肝脏、肾脏、精液以及眼睛的晶状体和角膜中EGT含量较丰富[3]。EGT可在分枝杆菌等微生物中合成,但人体无法自主合成EGT,需要通过饮食摄取并在体内积累。
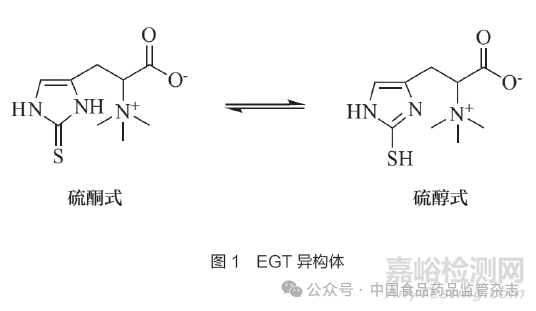
EGT化学名称为2-巯基组氨酸三甲基内盐,是一种手性小分子组氨酸衍生物,L-EGT为其天然存在形式。EGT纯品为白色无味晶体,相对分子质量229.3,易溶于水,在25℃水中的溶解度为0.9mol/L。EGT可结晶为二水合物或内盐,二水合物形式的EGT比无水内盐具有更强的吸湿性。EGT在水溶液中存在硫酮式和硫醇式2种异构体,如图1所示。在生理条件下,EGT主要以硫酮形式存在。EGT在生理和中等碱性(pH值为8~9)条件下都具有良好的稳定性,同时具有较高的热稳定性,其分解温度可达262~265℃[3-4]。EGT的标准氧化还原电位约为-0.06V,而其他天然硫醇抗氧化物质(如谷胱甘肽、半胱氨酸)的标准氧化还原电位一般为-0.32~-0.2V[5],表明EGT除具有良好的pH稳定性和热稳定性外,还表现出良好的抗氧化性。
作为一种亲水性两性离子化合物,EGT自身难以透过质膜,其在生物体内的运转依赖EGT转运体 (ergothioneine transporter,ETT,或称OCTN1,由基因SLC22A4编码)。ETT对EGT具有较高的亲和力,通过存在于细胞膜和线粒体膜上的ETT,EGT被转运至细胞中[6]。作为一种高效的转运体,ETT对EGT具有高度特异性[7]。研究显示,从斑马鱼中鉴定并敲除ETT后,EGT含量比野生型大幅降低[8]。只有表达ETT水平较高的细胞才能积累高水平EGT。在人体内,EGT的吸收、分配和保留完全取决于ETT的功能。缺乏ETT的细胞更容易受到氧化应激的影响,进而导致线粒体DNA损伤、蛋白质氧化和脂质过氧化增加[9-10]。
1、EGT的生物学功能
1.1 抗氧化
EGT是一种天然高效的抗氧化剂,其在体外实验中被证实对羟基自由基、次氯酸和过氧亚硝酸盐具有强大的清除能力。与谷胱甘肽等其他硫醇化合物相比,EGT能够以更高的速率灭活单线态氧[3];与尿酸和奎诺二甲基丙烯酸酯等抗氧化剂相比,EGT对过氧自由基的抗氧化能力高于奎诺二甲基丙烯酸酯,对羟基自由基和过氧亚硝酸盐的清除能力高于尿酸[11]。
在机体内,EGT是一种保护受损组织的适应性抗氧化剂。EGT在体内的分布表明,该化合物更倾向于积累在肝脏、红细胞、眼睛晶状体和精液等氧化应激和炎症水平较高的器官、细胞和分泌物中。在健康机体中,EGT的抗氧化作用可能有限,但当组织损伤、活性氧和活性氮过多或涉及氧化损伤水平增加时,EGT则表现出显著的保护作用[12]。氧化应激是机体在遭受有害刺激时,体内自由基过多产生,氧化还原系统动态失衡,从而导致机体受到损伤的过程。作为细胞抗氧化防御系统的一部分,Kelch样环氧氯丙烷相关蛋白1(Kelch-like ECH-associated protein 1,Keap1)-核因子E2相关因子2 (nuclear factor erythroid 2-related factor 2,Nrf2)/抗氧化反应元件 (antioxidant response element,ARE)信号通路在维持氧化还原平衡、防御氧化应激和炎症方面发挥着关键作用。在正常情况下,Nrf2与Cullin3(CUL3)和Keap1结合;当暴露于氧化应激时,Keap1/CUL3的多泛素化受阻,导致Nrf2释放并易位到细胞核,与ARE结合,触发多种抗氧化基因的激活[13]。EGT的抗氧化作用与Nrf2/ARE介导的细胞抗氧化基因激活密切相关,EGT促进Nrf2的核易位和激活,从而介导血红素加氧酶-1(heme oxygenase-1,HO-1)、NAD(P)H醌氧化还原酶1[NAD(P)H quinone oxidoreductase 1,NQO1]和γ-谷氨酸半胱氨酸连接酶(glutamate-cysteine ligase catalytic,GCLC)基因的表达。研究证明,Nrf2易位由磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)/蛋白激酶 B(protein kinase B,AKT)、蛋白激酶 C(protein kinase C,PKC)或活性氧信号级联介导[14-15]。此外,EGT还通过其抗氧化活性抑制核因子κB(nuclear factor-κB,NF-κB)转录途径,保护大鼠嗜铬细胞瘤细胞(PC12细胞)免受高血糖损伤[16],调节过氧化氢诱导的PC12细胞DNA损伤、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)激活和细胞死亡[17]。
EGT可降低小鼠运动后肌肉中代谢应激和炎症标志物以及氧化损伤标志物的表达,对延长运动力竭时间和运动后肌肉恢复有重要作用[18]。EGT也是二价金属特别是具有氧化还原活性的铜和铁的优秀螯合剂,通过形成非氧化还原活性的EGT-铜复合物,有效防止铜诱导的DNA和蛋白质的氧化损伤[19]。此外,EGT还可以减少亚硝化应激对乳铁蛋白的影响,从而维持其蛋白质活性[20]。
1.2 免疫调节
EGT具有免疫调节特性,可增强toll样受体(toll-like receptor,TLR)信号转导,用EGT预处理巨噬细胞,促进TLR配体诱导的细胞因子产生,并增强CD4+T细胞的辅助性T细胞17(Th17)极化[21]。EGT与TLR2配体共同作用,能够调节抑制微环境,增强免疫功能,降低肿瘤相关巨噬细胞的免疫抑制功能。TLR2刺激协同EGT可下调程序性细胞死亡蛋白配体1、集落刺激因子1受体、精氨酸酶1、脂肪酸合酶配体和肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)相关凋亡诱导配体在肿瘤相关巨噬细胞中的表达[22]。
EGT显著增加了神经祖细胞神经元标志物βⅢ-微管蛋白的免疫反应细胞数量,降低了星形胶质细胞标志物胶质原纤维酸性蛋白的免疫反应细胞数量,同时上调了神经元分化激活因子基因Math1的表达[23]。此外,EGT通过下调CD4+T细胞和巨噬细胞的数量,稳定了葡聚糖硫酸钠引起的免疫应答,可缓解其诱导的结肠炎[24]。
1.3 抗炎
炎症是人体应对各种损伤因子的刺激所发生的以防御反应为主的基本病理过程。研究表明,EGT能够抑制过氧化氢和TNF-α介导的NF-κB 激活,减少肺泡上皮细胞白细胞介素(interleukin,IL)-8释放,从而抑制慢性炎症性肺病的炎症反应[25]。EGT口服后会被机体吸收和保留,血浆和全血EGT水平显著升高,氧化损伤和炎症的生物标志物如8-羟基-2'-脱氧鸟苷、8-异前列腺素F2α和C反应蛋白等呈下降趋势[26]。
EGT具有调节NF-κB、MAPK和PI3K/AKT信号通路的作用,可显著降低TNF-α和IL-6水平及半胱氨酸天冬氨酸蛋白酶-3(cysteinyl aspartate specific proteinase-3,caspase-3)活性,抑制铁诱发的炎症产生,减少铁引起的肝细胞凋亡[27]。此外,EGT还抑制了TNF-α诱导的脂肪细胞炎症反应[28]。通过激活骨关节炎软骨细胞中的沉默信息调节因子6(silent information regulator 6,SIRT6)通路,EGT能够抑制NF-κB活性,从而显著减弱IL-1β引起的炎症反应,对骨关节炎的发展具有抑制作用[29]。在妊娠糖尿病患者中,EGT可改善NOD样受体热蛋白结构域相关蛋白 3(NOD-like receptor pyrin domain-containing protein 3,NLRP3)诱导的、由巨噬细胞亚群产生的线粒体超氧化物升高引发的胎盘炎症[30]。在心血管疾病和糖尿病患者中,内皮细胞暴露于高水平的胆固醇氧化产物7-酮胆固醇会介导病理性炎症,EGT可以显著抑制7-酮胆固醇诱导的促炎细胞因子IL-1β、IL-6、IL-8、TNF-α和环氧合酶-2的mRNA表达及其酶活性的升高[31]。
1.4 抗衰老
EGT对一些与衰老有关的疾病具有保护和预防作用。端粒缩短与衰老及多种年龄相关的疾病有关,而氧化应激通常是端粒缩短的主要原因之一。EGT在氧化应激条件下,通过降低端粒缩短率和保持端粒长度,显示出其有益的抗衰老作用[32]。
EGT有助于延缓衰老期间的运动功能衰退[33]。研究表明,EGT可以延长果蝇的寿命及攀爬能力。其延长寿命的潜在机制可能包括3个方面:通过协调胆碱能神经传递、酪氨酸代谢和过氧化物酶体蛋白来维持中枢神经系统的正常状态;通过改变组织蛋白酶D来调节自噬活性;以及通过控制底物进入三羧酸循环来维持正常的线粒体功能[34]。
每天摄入4~5mg/kg的EGT也被证明可显著延长雄性小鼠的寿命[35]。EGT还能够通过调节SIRT1和SIRT6信号通路,防止与高血糖相关的内皮细胞衰老[36]。此外,富含EGT的香菇提取物则能逆转叔丁基过氧化氢诱导的神经元细胞衰老[37]。
1.5 皮肤损伤修复
皮肤暴露在太阳紫外线照射下会导致DNA、蛋白质和脂质损伤,最终导致皮肤光老化。EGT积累在表皮角质细胞中,通过降低活性氧水平、保持细胞活力和消除大规模凋亡反应来保护角质细胞免受紫外线损害[38]。
用EGT预处理角质形成细胞可显著抑制Nrf2/HO-1通路和热休克蛋白70的下调,通过抑制活性氧的产生以及抑制caspase-8和聚腺苷二磷酸核糖聚合酶的裂解保护角质细胞,显著降低了旁分泌细胞因子IL-1β、IL-6和TNF-α的水平[39]。EGT可提高Hs68人皮肤成纤维细胞受紫外线照射后的细胞活力,抑制紫外线照射后Hs68细胞中活性氧的产生,同时表现出对基质金属蛋白酶-1的抑制作用和对I型原胶原的恢复效果[40]。此外,EGT还被证明有助于缺血性皮肤伤口的愈合[41]。
1.6 神经系统调节
EGT缺乏可能会增加个体患神经退行性疾病的风险。研究发现,老年人的全血EGT水平显著下降,患轻度认知障碍的受试者血浆EGT水平也显著低于同龄健康受试者[42]。服用EGT可增强记忆功能,防止压力引起的睡眠障碍和神经元损伤。EGT通过其抗氧化活性以及促进神经发生和神经元成熟,有益于大脑功能[43]。服用EGT还可能具有抗抑郁作用[44],防止应激引起的睡眠障碍,尤其是与抑郁症相关的睡眠障碍[45]。
促进神经发生被认为是治疗神经精神疾病的有效策略之一。EGT可通过核糖体蛋白S6激酶1在Thr389位点的磷酸化以及激活原肌球蛋白受体激酶 B信号传导,介导神经干细胞的神经元分化[46]。通过其抗氧化活性,EGT对HT22海马神经元具有神经保护作用[47]。在阿尔茨海默病的研究中,β淀粉样蛋白被认为与疾病的发生和进展相关。通过抑制过氧亚硝酸盐的形成和蛋白质酪氨酸残基的硝化作用,EGT有效抑制了β淀粉样蛋白诱导的PC12细胞凋亡,在阿尔茨海默病和其他神经退行性疾病的治疗中起到神经保护的作用[48]。此外,EGT通过发挥抗氧化作用,可防止6-羟基多巴胺引起的神经元细胞死亡,这可能是预防帕金森病发生发展的有效方法[49]。
2、EGT的制备
EGT的制备方法主要包括天然产物提取法、化学合成法和生物合成法3种。
2.1 天然产物提取法
天然产物如食用菌、麦角、谷物及动物组织中含有EGT,其中食用菌EGT含量较高,是EGT的重要来源。例如,牛肝菌中EGT含量为1812μg/g,杏鲍菇中EGT含量为542μg/g,香菇中EGT含量为353μg/g,黑木耳中EGT含量为94μg/g[50]。Zhu等[51]研究了不同营养和环境条件对紫革耳(Panus conchatus)生长和EGT产量的影响,发现糖蜜和大豆蛋白胨能促进细胞生长及EGT的积累,添加组氨酸、甲硫氨酸和半胱氨酸前体均能提高EGT的产量,获得的EGT产量达148.79mg/L。灵芝作为一种传统的药用菌类,含有较高浓度的EGT。Xie等[52]采用灵芝原生质体融合技术,获得的融合菌株中EGT产量达12.70mg/L±1.85mg/L,显著高于亲本菌株。
天然产物提取法一般包括回流提取法、酶解提取法、超声微波联合法等。Zhang等[53]评价了不同干燥工艺对金顶侧耳(Pleurotus citrinopileatus)中EGT含量的影响,采用高静压萃取法提取EGT,用响应面分析法对提取参数进行优化。结果表明,自然通风干燥法可增加蘑菇的EGT含量,最佳提取条件为:压力250MPa、提取时间52min、蒸馏水为提取溶剂、液固比为1:10,得到的EGT含量最高,为4.03mg/g±0.01mg/g。
由于天然产物的EGT含量普遍很低,提取时需要耗费大量原料,提取成本高昂,同时也存在原料杂质多、药物残留等问题,这些因素限制了天然产物提取法的产业化应用。
2.2 化学合成法
传统的EGT化学合成法以组氨酸为起始原料,通过巯基化、巯基保护、甲基化、脱保护等多步反应在水溶液中合成EGT,合成路线冗长且收率较低[54-55]。近年来,有研究模仿生物合成路径,改进了EGT的化学合成法。例如,Erdelmeier等[56]以组氨酸甜菜碱、半胱氨酸、巯基丙酸等为原料,用简化的一锅反应制得EGT,避免引入巯基保护基团,缩短了合成路线并提高了收率。EGT仿生化学合成法见图2。
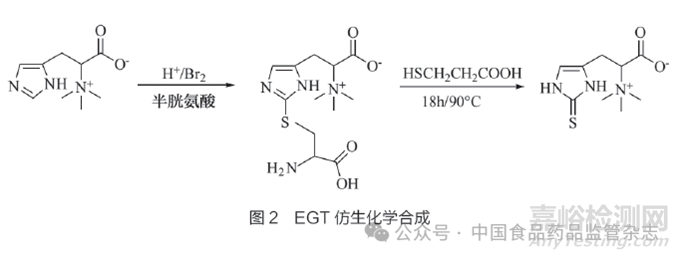
化学合成法也存在一些挑战。例如,羧基α位碳的手性在酸碱条件下易发生消旋,这使得高光学纯度EGT不易获得。此外,化学合成法所需原料价格较高,原料2-巯基咪唑不易制备[55],产物后处理也比较困难。因此,研究者需要对化学合成法进行优化改进,以满足规模化工业生产的需求。
2.3 生物合成法
通过生物合成法获得EGT具有原料易获得、成本相对较低等优势,已成为近年来EGT的主要制备方式。在自然界中,EGT存在好氧生物合成和厌氧生物合成2种途径。
2.3.1 EGT好氧生物合成
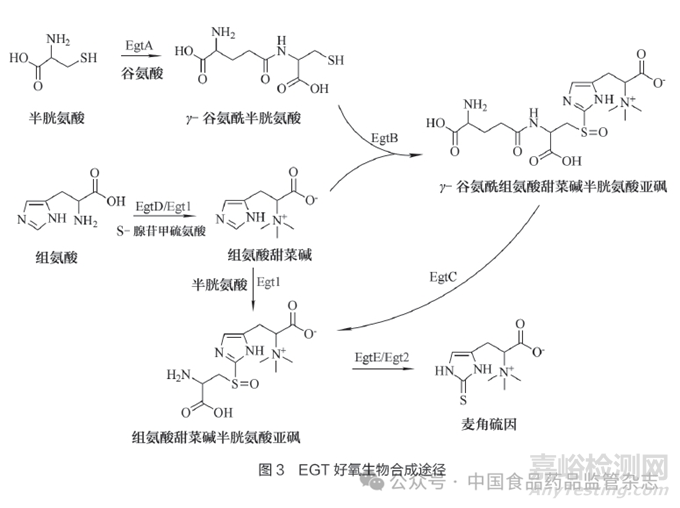
许多研究已阐明EGT的生物合成过程,基本上EGT好氧生物合成途径可以分为细菌途径和真菌途径。Seebeck等[57]在耻垢分枝杆菌(Mycobacterium smegmatis)中阐明并在体外重构了EGT的好氧生物合成途径,确定了合成途径涉及的5个基因EgtA、EgtB、EgtC、EgtD和EgtE,分别编码谷氨酰半胱氨酸合成酶(EgtA)、Fe2+依赖的氧化酶(EgtB)、谷氨酰转移酶(EgtC)、S-腺苷甲硫氨酸(S-adenosyl methionine,SAM)依赖的组氨酸甲基转移酶(EgtD)和 磷酸吡哆醛依赖的C-S裂解酶(EgtE)。其中,EgtA催化谷氨酸和半胱氨酸缩合生成γ-谷氨酰半胱氨酸;EgtB在氧气和硫酸亚铁存在下催化氧化偶联,将γ-谷氨酰半胱氨酸上的硫醇基添加到组氨酸甜菜碱(由EgtD催化组氨酸和SAM合成)的侧链上;再经EgtC的催化进一步形成组氨酸甜菜碱半胱氨酸亚砜;最后经EgtE催化脱去丙酮酸和氨生成EGT[58]。
EGT在真菌的生物合成途径中,主要涉及2个合成酶基因Egt1和Egt2。Bello等[59]鉴定了粗糙脉孢菌(Neurospora crassa)中一种特异性合成酶NcEgt1,NcEgt1包含类似于耻垢分枝杆菌 EgtB和EgtD中的结构域[60]。Egt1参与了2步催化反应,首先催化组氨酸转化为组氨酸甜菜碱,然后催化组氨酸甜菜碱与半胱氨酸结合形成组氨酸甜菜碱半胱氨酸亚砜。Egt2与EgtE类似,催化组氨酸甜菜碱半胱氨酸亚砜形成EGT[61]。如图3所示,由于只有2种酶参与,EGT在真菌中的生物合成途径比在细菌中的生物合成途径要简单得多。
2.3.2 EGT厌氧生物合成
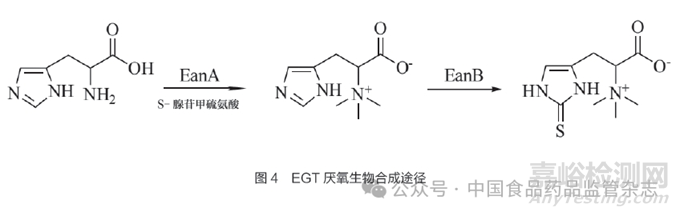
EGT的厌氧生物合成途径在一种严格厌氧的泥生绿菌(Chlorobium limicola)中有报道,其中EGT的生物合成是通过不依赖氧气的2步反应完成的。Burn等[62]从这种厌氧泥生绿菌中鉴定出了甲基转移酶 (EanA)和硫转移酶(EanB)。如图4所示,EanA 催化组氨酸转化为组氨酸甜菜碱,随后EanB在无氧条件下将硫原子转移至组氨酸甜菜碱的咪唑环生成EGT。尽管厌氧生物合成途径更为简单,但EanA和EanB的催化速率远低于好氧生物合成途径中EgtD和EgtB的催化速率。因此,目前EGT生物合成仍然以好氧生物合成途径为主。
基于EGT的生物合成途径,研究者构建了一系列工程菌株并优化其工艺条件,以提高EGT的生产效率。表1列举了部分EGT生物合成的研究成果及生产水平。

3、EGT的应用
3.1 EGT在疾病诊疗中的应用
EGT已被证实具有细胞保护作用,这使得EGT在慢性疾病如心血管和代谢疾病中具有潜在的治疗作用。这些慢性疾病一般都伴随持续的氧化应激和炎症。较高的EGT水平是心脏代谢疾病和死亡风险较低的独立标志物[72]。EGT可中断动脉粥样硬化相关的黏附分子表达的促炎诱导,预防动脉粥样硬化,降低心血管疾病风险[73]。EGT在糖尿病诊疗中也显示出重要意义。研究表明,给糖尿病大鼠在妊娠前11.5天每天补充EGT可降低胚胎畸形率[74]。EGT与二甲双胍联合给药可降低2型糖尿病大鼠的高血糖,抑制肾脏炎症并改善肾功能,这种疗效与EGT激活Nrf2抗氧化信号增强抗氧化防御系统有关[75]。
EGT在治疗先兆子痫中具有潜在应用。先兆子痫是一种基于胎盘异常的妊娠多因素性高血压疾病,人体内EGT水平与先兆子痫风险有显著的非线性关联[76]。EGT可改善子宫灌注压降低先兆子痫大鼠的高血压和胎儿体重,改善其肾脏和胎盘组织的线粒体功能[77]。
顺铂是一种有效的化疗药物,但其治疗应用受到肾毒性的限制。顺铂引起的肾毒性很大程度上归因于氧化应激的诱导和炎症反应。EGT通过靶向Nrf2、NF-κB、凋亡信号和抑制γ-谷氨酰转肽酶,可以减轻顺铂引起的肾毒性[78]。EGT还可通过抑制氧化应激和恢复神经细胞中的胆碱能神经活性来保护顺铂诱导的神经元损伤并增强认知能力[79]。
EGT也显示出预防阿尔茨海默病的潜力[80]。在包括伴有海马萎缩的阿尔茨海默病在内的痴呆症患者中,认知能力的恶化与EGT水平的下降有关,而硫酸吲哚酚和喹啉酸等化合物则增加,利用这些痴呆症标志物,可以进行诊断和治疗干预,以减缓认知能力下降[81]。
基于EGT可调节炎症、清除自由基、防止神经元损伤、防止急性呼吸窘迫综合征[82]等特性,也有学者提出了将EGT应用于治疗新型冠状病毒感染的可能性和前景[83]。
3.2 EGT新药研发进展
现有EGT新药的研究大多处于临床前研究阶段。毒理学研究结果显示,EGT耐受性良好,对所评估的生殖参数无不良影响[84];在测试条件下,对细菌测试菌株没有诱变活性[85]。浓度为5000μg/ml的EGT在CHL细胞染色体畸变试验中未引起结构性染色体畸变,在哺乳动物体内红细胞微核试验中,雄性小鼠口服1500mg/kg的EGT剂量,未观察到微核多染红细胞率增加[86]。在这些测试中,EGT未显示出潜在的遗传毒性、致癌性和致突变性。
EGT对中枢神经系统神经元具有保护作用。在啮齿动物模型中,脑室给药EGT可有效减少中风后的脑梗死体积[87]。EGT在口服给药后能促进正常小鼠海马区的神经元成熟,提高学习和记忆能力[88]。在治疗急性肺损伤方面,研究显示100mg/kg EGT与参照药物非布司他一样有效,临床试验后或可成为治疗该疾病的替代药物[89]。此外,EGT还是治疗胱氨酸结石[90]、预防结直肠癌[91]、治疗年龄相关性黄斑变性[92]的新型方法和潜在手段。
目前,已有一些关于EGT在体内吸收、分布与积累及其代谢的相关研究[93]。此外,还开展了一些关于EGT潜在治疗功效的临床研究。例如,一项近期研究显示,有高度焦虑和睡眠问题的受试者持续4周每天服用20mg EGT,睡眠困难显著改善[94]。另一项临床研究表明,EGT可以作为认知能力下降的预测性生物标志物[95]。
3.3 EGT在食品领域的应用
欧洲食品安全局(European Food Safety Authority,EFSA)已评估并声明,EGT用作婴幼儿、孕妇和哺乳期妇女营养补充剂是安全的[96]。蘑菇是EGT的主要食用来源,食用含EGT的蘑菇后,EGT可被红细胞吸收并减弱餐后的甘油三酯应答[97]。增加蘑菇的摄入可以降低慢性衰老疾病的发病率[98]。
EGT对肉类颜色的稳定和保质期延长具有重要作用。在香肠中添加富含EGT的金针菇提取物能有效预防脂质和蛋白质氧化[99]。EGT对金枪鱼肉中高铁肌红蛋白的形成具有抑制作用,在牛肉和鱼类中添加含EGT的蘑菇提取物,可以控制在冰冻储存过程中的脂质氧化,肉类颜色保持时间明显得以提升[100]。用富含EGT的壳聚糖涂层处理海鲈,可降低其腐胺、尸胺和组胺等生物胺含量,延长海鲈保质期[101]。此外,喷洒EGT水溶液可降低双孢菇(Agaricus bisporus)采后褐变的程度,从而延长保质期[102]。
EGT还是一种替代合成黑变病抑制剂的天然成分。含EGT的金针菇提取物可显著降低虾血淋巴多酚氧化酶活性和血细胞酚氧化酶原基因的表达,有效预防虾蟹在收获和储存期的黑变[103-104]。
3.4 EGT在化妆品领域的应用
EGT具有多种皮肤护理功能,已被国家药品监督管理局发布的《已使用化妆品原料目录(2021年版)》收录。EGT可应用于抗衰老化妆品。一方面,EGT可预防光老化,通过抑制髓过氧化物酶活性和清除卤化物来保护紫外线B段诱导的DNA损伤[105]。可见光通过类似于紫外线A段辐射诱导的氧化应激机制对人角质形成细胞和CHO细胞产生遗传毒性,EGT有效防止了可见光和紫外线A段辐照引起的DNA损伤[106]。另一方面,EGT可抑制基质金属蛋白酶-1的激活和I型原胶降解[14],具有对抗氧化应激、清除自由基的能力,通过Nrf2途径促进抗氧化剂的上调和螯合金属离子,在防止衰老过程中发挥着重要作用[107]。
基于良好的抗炎功能,EGT可用于皮肤舒缓类化妆品中,应对外界刺激而导致的皮肤炎症。EGT对酪氨酸酶活性也具有一定的抑制作用,是一种非竞争性酪氨酸酶抑制剂。这种抑制作用与其抗氧化活性相关性较弱,可能与其结构中存在的硫代咪唑环有关[108],这也意味着EGT在美白类化妆品中具有一定的应用潜力。
4、结 语
EGT已被证明具有抗氧化、抗炎、调节免疫等多种生物学功能,使其在药品、食品、化妆品等领域有着良好的应用前景。目前,EGT研究主要集中在生物合成和在疾病诊疗、食品、化妆品等领域的应用。在药品领域,虽然现有研究显示EGT具有多种潜在的治疗作用,但对其体内效应及药效发挥机制的研究尚不充分,临床试验也需进一步开展。蘑菇是EGT的主要食用来源,增加蘑菇摄入有益健康,但饮食中理想的EGT含量尚不明确。在化妆品领域,随着近年来天然成分和功效化妆品的兴起,EGT也成为备受瞩目的活性成分。然而,目前EGT相对昂贵的市场价格限制了其在化妆品领域的广泛使用,这需要研究者进一步优化EGT生产过程,寻求经济高效的生产方式。此外,作为皮肤保护剂,EGT在皮肤中的渗透和利用、皮肤保护作用机制以及EGT与其他护肤活性成分之间的协同效应也有待进一步研究和阐明。
引用本文
刘向前,邱晓锋,刘忠*.麦角硫因的功能、制备及应用研究进展[J].中国食品药品监管,2024(6):116-128.

来源:中国食品药品监管杂志


